
HOME > 試薬 > 生化学一般試薬/その他 > 蛍光/標識用試薬 > Spirochrome イメージング試薬
Spirochrome イメージング試薬
掲載日情報:2023/10/05 現在Webページ番号:69536

Spirochrome社ではライブセルイメージング、超解像顕微鏡、膜張力解析、膨張顕微鏡法(ExM)に対応したイメージング試薬を幅広く取りそろえています。細胞中のアクチン、DNA、チューブリン、RNA、ミトコンドリア、細胞膜などのライブイメージングや、膜張力の変化の可視化といった、観察対象や解析目的に応じて選択できます。
Cytoskeleton社では、2015年よりSpirochrome社製品を取り扱っています。
追加しました。
- さまざまなターゲットに対応するライブイメージングプローブ
- 長時間ライブセル観察に適した膜・ミトコンドリアプローブ
- ライブセルRNA蛍光イメージングプローブ
- 脂質二重膜の張力変化を可視化する機能解析プローブ
- 膨張顕微鏡法(ExM)でアクチンを高解像度に可視化するプローブ
- 反応性官能基を有する蛍光色素
さまざまなターゲットに対応するライブイメージングプローブ
SiR Probe/SPY Probe
DNAからオルガネラ、細胞骨格、タグ融合タンパク質など、さまざまなターゲットに対応する細胞透過性のライブセルイメージングプローブです。SiRプローブは、シリコンローダミン(Silicon Rhodamine:SiR)ベースの遠赤色のフルオロジェニック特性を有する蛍光プローブで、超解像顕微鏡法による解析にも使用できます。SPYプローブは、SiRプローブの優れた特性に加え、FITCやTMR、またはTexas Red用など異なるチャンネルで解析することができます。SiR Probe/SPY Probeの詳細はこちらをご覧下さい。
主な製品ラインナップ商品コード(#CY-SCXXX)をクリックすると各製品の価格表をご覧いただけます。
| シリーズ名 | SiRプローブ | SPYプローブ | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 蛍光色素 | SiR | SiR650 | SiR700 | SPY505 | SPY555 | SPY595 | SPY620 | SPY650 | SPY700 | ||
| 測定 波長 |
励起(nm) | 652 | 652 | 689 | 512 | 555 | 599 | 619 | 652 | 696 | |
| 蛍光(nm) | 674 | 674 | 716 | 531 | 580 | 615 | 636 | 674 | 718 | ||
| 標的 因子 |
DNA | CY-SC007 | - | CY-SC015 | CY-SC101 | CY-SC201 | CY-SC301 | CY-SC401 | CY-SC501 | CY-SC601 | |
| Actin | Actin | CY-SC001 | - | CY-SC013 | - | CY-SC202 | - | CY-SC402 | - | - | |
| FastAct | - | - | - | - | CY-SC205 | - | - | CY-SC505 | CY-SC605 | ||
| FastAct_X | - | - | - | - | CY-SC209*2 | - | - | CY-SC502 | - | ||
| Tubulin | CY-SC002 | - | CY-SC014 | - | CY-SC203 | - | - | CY-SC503 | - | ||
| Peroxisome | - | - | - | - | CY-SC207 | - | - | CY-SC507 | - | ||
| Golgi | - | - | - | - | CY-SC208 | - | - | CY-SC508 | - | ||
| Lysosome | CY-SC012 | - | CY-SC016 | - | - | - | - | - | - | ||
| Tag | SNAP Tag | - | CY-SC504 | CY-SC604*1 | - | CY-SC204 | - | CY-SC404 | - | - | |
| HaloTag*3 | CY-SC506 | - | - | CY-SC106 | CY-SC206 | - | - | - | CY-SC606 | ||
*1 SiR700-BG(#CY-SC604)の励起/蛍光波長は696/718 nmです。
*2 SPY555-FastAct_X(#CY-SC209)の励起/蛍光波長は564/589 nmです。
*3 HaloTag検出用シリーズにはSiR/SPYプローブ以外の製品もあります。詳細は、上記表中のシリーズ名からご確認いただけます。
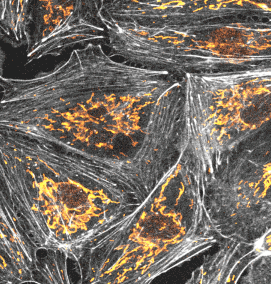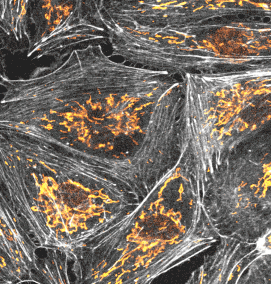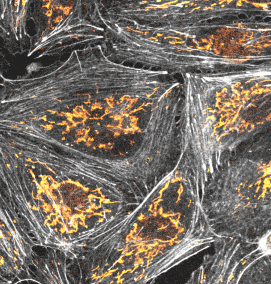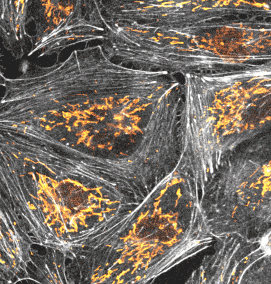
使用例
画像をクリックすると、拡大画像が表示されます。
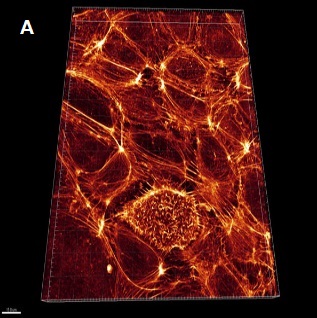
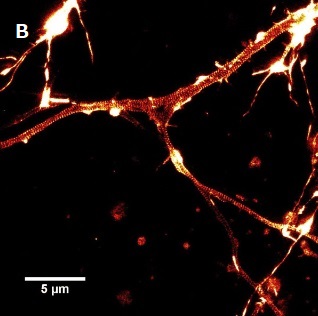
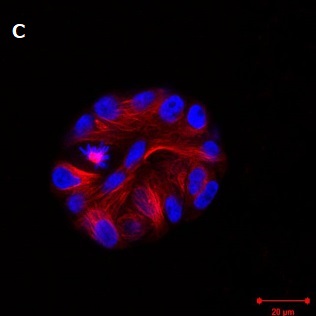
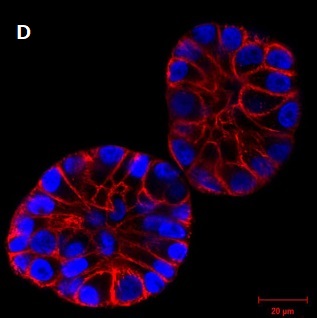
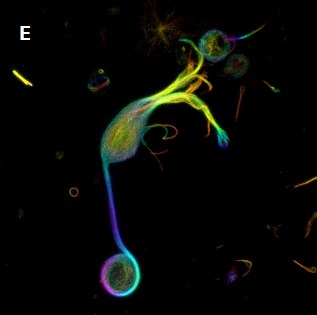
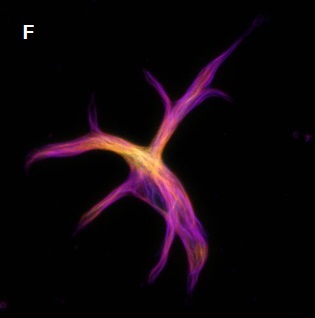
追加しました。
長時間ライブセル観察に適した膜・ミトコンドリアプローブ
PKmem Probe

PKmito Probe
追加しました。
ライブセルRNA蛍光イメージングプローブ
SPYRHO 555

追加しました。
脂質二重膜の張力変化を可視化する機能解析プローブ
Flipper-TRシリーズ
Flipper-TRシリーズは、それぞれ細胞の原形質膜、および各種のオルガネラ(小胞、リソソーム、ミトコンドリア)の脂質二重層に特異的に挿入され、蛍光寿命の変化によりそれぞれの膜の張力変化を検出できる蛍光プローブです。これらの4種類の膜プローブは、同一の励起・蛍光波長(Ex 488 nm/Em 575~625 nm)を用いた多重標識実験にも適応します。生細胞の張力をリアルタイムで測定できます。Flipper-TRシリーズの詳細はこちらをご覧下さい。
また、関連製品としてHaloTag融合タンパク質を特異的に標識し、蛍光寿命の変化を通じて膜張力の変化を検出する蛍光プローブHalo-Flipperもあります。
追加しました。
膨張顕微鏡法(ExM)でアクチンを高解像度に可視化するプローブ
HAK-actin

追加しました。
反応性官能基を有する蛍光色素
タンパク質や生体分子の標識に使用できるアミン反応性、チオール反応性(マレイミド)、カルボン酸修飾の色素に加え、クリックケミストリーに有用なアジド、テトラジン修飾色素があります。
商品コードをクリックすると各製品の価格表をご覧いただけます。
| 品名 | SiR-Azide | SiR-BCN | SiR-COOH | SiR-NHS | SiR-Tetrazine | |
|---|---|---|---|---|---|---|
| 修飾基 | Azide | BCN | -COOH | -NHS | Tetrazine | |
| 蛍光波長 (nm) |
励起 | 652 | ||||
| 蛍光 | 674 | |||||
| 商品コード | CY-SC009 | CY-SC017 | CY-SC004 | CY-SC003 | CY-SC008 | |
追加しました。
製品情報は掲載時点のものですが、価格表内の価格については随時最新のものに更新されます。お問い合わせいただくタイミングにより製品情報・価格などは変更されている場合があります。
表示価格に、消費税等は含まれていません。一部価格が予告なく変更される場合がありますので、あらかじめご了承下さい。