
HOME
>
試薬
>
アポトーシス
>
アポトーシス誘導/阻害物質
>
ニューロン用リプログラミング関連研究用化合物
HOME
>
試薬
>
天然物/有機化合物/糖質
>
天然物/有機化合物
>
生理活性物質
>
ニューロン用リプログラミング関連研究用化合物
神経細胞のリプログラミングに有用な低分子化合物(Cayman Chemical社) ニューロン用リプログラミング関連研究用化合物
掲載日情報:2020/02/13 現在Webページ番号:69240

Cayman Chemical社では、ニューロン用リプログラミング関連研究用化合物を多数取りそろえています。低分子化合物を組み合わせることにより、遺伝子導入せずにマウスの線維芽細胞を機能性ニューロン、神経前駆細胞、または神経幹細胞ニューロンへとリプログラミングすることができます。また、クロマチンを修飾することや幹細胞の自己複製を促進することも示されています。
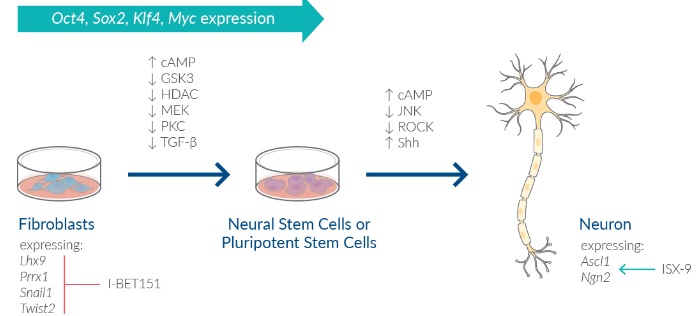
化学的に誘導した繊維芽細胞(fibroblast)から幹細胞(stem cells)とニューロン(neurons)へのリプログラミング
低分子を組み合わせることにより、マウス線維芽細胞を機能性ニューロン、神経前駆細胞、神経幹細胞に変換できます。これらの化学物質の多くは、同じ標的に対する別の阻害物質と交換でき、同じ効果が得られます。
追加しました。
エピジェネティック修飾因子
エピジェネティック修飾因子とリプログラミング
低分子エピジェネティック修飾因子には、以下のようなエピジェネティック修飾機能が含まれます。
- DNAの過剰なメチル化:転写因子のプロモーターへの結合阻害による遺伝子の下方制御とサイレンシングに関与しています。
- メチルトランスフェラーゼ阻害物質:高メチル化多能性遺伝子を再活性化します。
- ヒストンの脱アセチル化:クロマチン構造の変化による遺伝子発現、細胞周期の進行、細胞の生存・分化に関与しています。
- HDAC阻害物質:HDAC阻害物質を用いることで、Oct4とSox2の発現に加えて多能性遺伝子を活性化し、体細胞リプログラミング誘導プロトコルでのc-MycおよびKlf4がん遺伝子が不要になります。この方法は、成体細胞にOct4、Sox2、Klf4、c-Mycの4つの遺伝子を導入するだけで多能性幹細胞が得られることを示した山中伸弥教授らの方法でのリプログラミング効率を2倍に向上させることができます。また、HDAC阻害物質は、細胞増殖の増加、アポトーシスの阻害、G2/M期の遮断を防止することにより、細胞を老化ストレスから保護すると考えられています。
- BRD BETブロモドメインの阻害:線維芽細胞状態の抑制に必要なコア転写ネットワークを破壊します。
製品ラインナップ
品名/構造式をクリックするとメーカーサイトでの製品の詳細、商品コードをクリックすると各製品の価格表をご覧いただけます。
| 品名/構造式 | 商品コード | CAS番号 | 化学式 | M.W. | 純度 | 標的因子 |
|---|---|---|---|---|---|---|
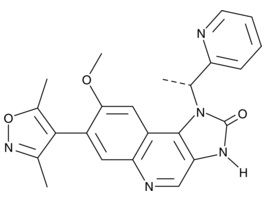
| I-BET151
|
11181 | 1300031-49-5 | C23H21N5O3 | 415.5 | ≧98% | BRD2, BRD3, BRD4 |
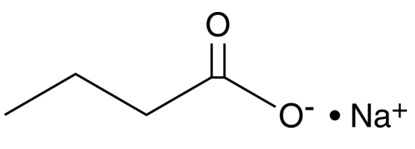
| Sodium Butyrate
|
13121 | 156-54-7 | C4H7O2・Na | 110.1 | ≧95% | HDACs |
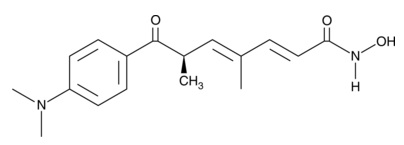
| Trichostatin A
|
89730 | 58880-19-6 | C17H22N2O3 | 302.4 | ≧98% | |
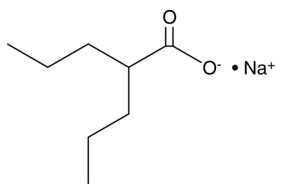
| Valproic Acid, Sodium Salt
|
13033 | 1069-66-5 | C8H15O2・Na | 166.2 | ≧95% | |
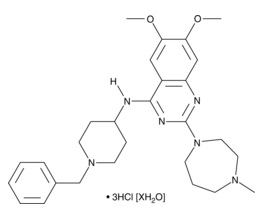
| BIX01294 (hydrochloride hydrate)
|
13124 | 1808255-64-2 | C28H38N6O2・3HCl [XH2O] | 600.0 | ≧98% | G9a histone methyltransferase |
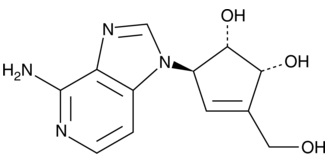
| 3-Deazaneplanocin A
|
13828 | 102052-95-9 | C12H14N4O3 | 262.3 | ≧97% | EZH2 lysine methyltransferase |
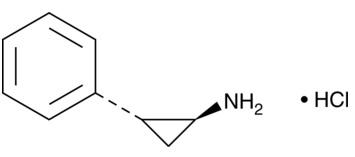
| 2-PCPA
|
10010494 | 1986-47-6 | C9H11N・HCl | 169.7 | ≧98% | Lysine-specific demethylase 1 |
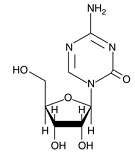
| 5-Azacytidine
|
11164 | 320-67-2 | C8H12N4O5 | 244.2 | ≧95% | DNA methyltransferases |
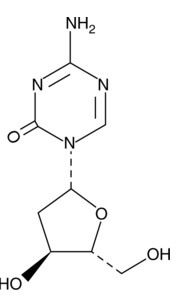
| Decitabine
|
11166 | 2353-33-5 | C8H12N4O4 | 228.2 | ≧98% | |
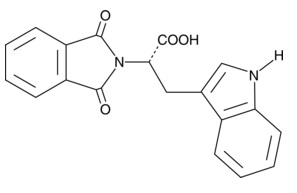
| RG-108
|
13302 | 48208-26-0 | C19H14N2O4 | 334.3 | ≧98% |
追加しました。
細胞シグナル伝達経路とアポトーシスの修飾因子
MEMO 細胞シグナル伝達経路とアポトーシスの修飾因子とは
- Rhoキナーゼ(ROCK)シグナル伝達経路の阻害:多能性幹細胞、およびニューロンの生存の維持、リプログラミング効率の改善します。
- Wntシグナル伝達とGSK3活性の制御:安静状態で、Wntシグナル伝達はβ-カテニンのGSK3リン酸化により阻害され、β-カテニンを不安定化し分解します。GSK3阻害物質は、多能性細胞からの神経発達を誘導します。
- MEK阻害物質:多能性を促進し、非特異的な分化を誘導します。
- GSK3:40以上のタンパク質標的があり、リプログラミングにも影響を与える可能性のある代謝調節を含む他のシグナル伝達経路にも関与しています。
細胞シグナル伝達経路とアポトーシスの修飾因子製品ラインナップ
品名/構造式をクリックするとメーカーサイトでの製品の詳細、商品コードをクリックすると各製品の価格表をご覧いただけます。
| 品名/構造式 | 商品コード | CAS番号 | 化学式 | M.W. | 純度 | 標的因子 |
|---|---|---|---|---|---|---|
| Thiazovivin
|
14245 | 1226056-71-8 | C15H13N5OS | 311.4 | ≧98% | ROCK |
| Y-27632 (hydrochloride)
|
10005583 | 129830-38-2 | C14H21N3O・2HCl | 320.3 | ≧98% | |
| CHIR99021
|
13122 | 252917-06-9 | C22H18Cl2N8 | 465.3 | ≧95% | GSK3α, GSK3β |
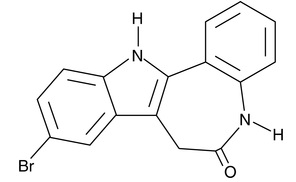
| Kenpaullone
|
10010239 | 142273-20-9 | C16H11BrN2O | 327.2 | ≧98% | GSK3β |
| PD 0325901
|
13034 | 391210-10-9 | C16H14F3IN2O4 | 482.2 | ≧98% | MEK |
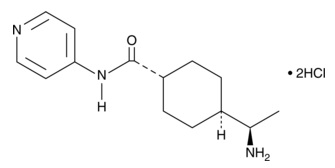
| A 83-01
|
9001799 | 909910-43-6 | C25H19N5S | 421.5 | ≧95% | ALK4, ALK5, ALK7 |
| ALK5 Inhibitor II (RepSox)
|
14794 | 446859-33-2 | C17H13N5 | 287.3 | ≧95% | ALK5 |
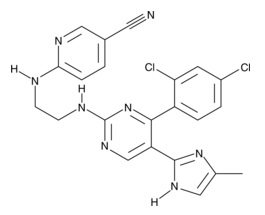
| LDN-193189
|
11802 | 1062368-24-4 | C25H22N6 | 406.5 | ≧95% | ALK1, ALK2, ALK3, ALK6 |
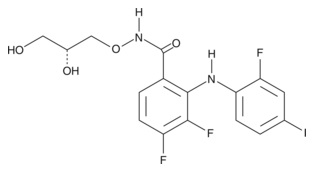
| SB-431542 (hydrate)
|
13031 | - | C22H16N4O3・XH2O | 384.4 | ≧98% | ALK4, ALK5, ALK7 |
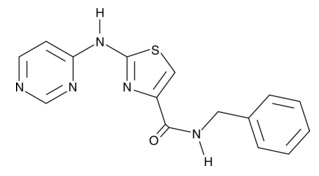
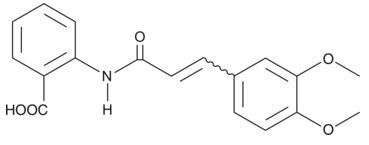
| Tranilast
|
13044 | 53902-12-8 | C18H17NO5 | 327.3 | ≧98% | TGF-β1 |
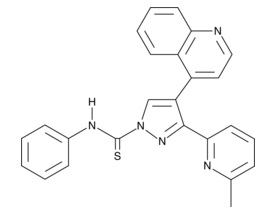
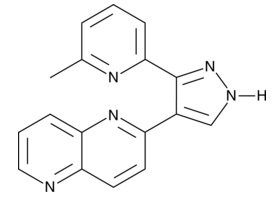
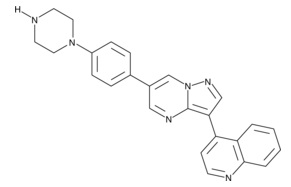
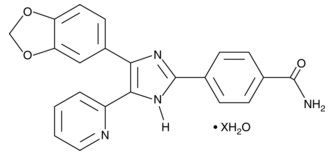
追加しました。
その他のリプログラミング制御因子
MEMO その他のリプログラミング制御因子とリプログラミング
- Smoothend(SMO):Shhシグナル伝達を媒介する細胞表面レセプターで、SMOアゴニストは神経幹細胞の分化の促進に用いられます。
- アデニルシクラーゼ(またはcAMP分解阻害物質):セカンドメッセンジャーのcAMPレベルを増加させます。また、cAMPの下流にあるCREB(cAMP-responsive element binding)は、ニューロンの分化を増強します。cAMP活性化物質は、Oct4の代替物質となります。
- c-Jun N-terminal kinases (JNKs):ストレスとアポトーシス、細胞運命の決定と分化への適応を媒介する構成要素。他のシグナル伝達阻害物質と組み合わせることにより、線維芽細胞の神経変換の促進、およびG2/M期の細胞を停止します。
- プロテインキナーゼC(PKC):胚性幹細胞の自己複製と系統のコミットメントのバランスをとる上で重要であり、神経細胞系統の形成中の遺伝子発現の調節に関与している可能性があります。PKC阻害物質は、線維芽細胞の多能性状態へのリプログラミングを可能にし、神経変換を促進します。
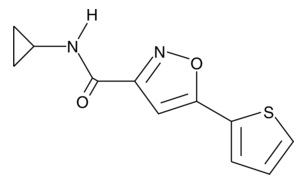
- イソオキサゾール9(ISX-9):神経分化の促進、筋細胞エンハンサー因子2(MEF2)の発現に関わっています。
- MEF2:初期発生において前ニューロン因子および生存促進因子として重要な転写因子です。
- 活性型レチノイド:レチノイドXレセプター(RXR)に関連し、核内のretinoic acid DNA response elements に結合するレチノイド酸レセプター(RAR)転写因子に結合し、分化を誘導します。レチノイドによる結合は、RAR/RXRコリプレッサーとコアアクティベータータンパク質間の相互作用の変化を開始し、主要な標的遺伝子の転写を活性化します。また、エピジェネティックな変化を誘発するタンパク質との相互作用を変更し、転写因子をコードする遺伝子の転写とさらなる遺伝子発現を修飾するタンパク質のシグナル伝達します。さらにレチノイドがレチノイン酸非依存性機構を介し、自己再生のための内因性機構を活性化することにより、幹細胞の自己再生を支援することが示されています。
その他のリプログラミング制御因子製品ラインナップ
品名/構造式をクリックするとメーカーサイトでの製品の詳細、商品コードをクリックすると各製品の価格表をご覧いただけます。
| 品名/構造式 | 商品コード | CAS番号 | 化学式 | M.W. | 純度 | 標的因子 |
|---|---|---|---|---|---|---|
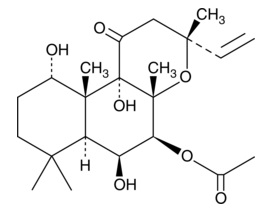
| 7-keto-25-hydroxy Cholesterol
|
25973 | 64907-23-9 | C27H44O3 | 416.6 | ≧95% | SMO |
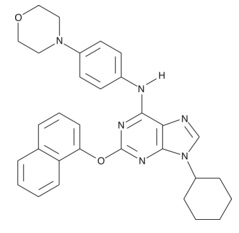
| Purmorphamine
|
10009634 | 483367-10-8 | C31H32N6O2 | 520.6 | ≧98% | |
| Forskolin
|
11018 | 66575-29-9 | C22H34O7 | 410.5 | ≧98% | cAMP |
| SP 600125
|
10010466 | 129-56-6 | C14H8N2O | 220.2 | ≧98% | JNK |
| Gö 6983
|
13311 | 133053-19-7 | C26H26N4O3 | 442.5 | ≧98% | PKC |
| ISX-9
|
16165 | 832115-62-5 | C11H10N2O2S | 234.3 | ≧95% | MEF2-dependent signaling |
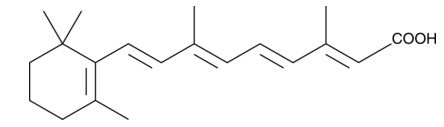
| all-trans Retinoic Acid
|
11017 | 302-79-4 | C20H28O2 | 300.4 | ≧98% | RARs |
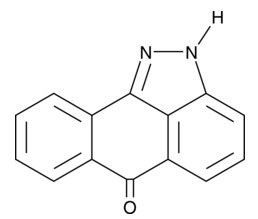
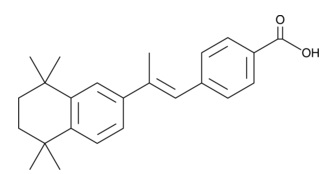
| TTNPB
|
16144 | 71441-28-6 | C24H28O2 | 348.5 | ≧98% |


追加しました。
細胞老化の減速物質
MEMO p53の役割
p53の活性化は、細胞周期をブロックする転写イベントを誘発し、DNAの修復、老化、またはアポトーシスを介して細胞の恒常性を回復します。この経路は、がん細胞の増殖防止に重要ですが、老化にも関与し、早期の幹細胞の枯渇に関連します。p53は、DNA損傷後の胚性幹細胞の多能性を抑制し、体細胞の人工多能性幹細胞へのリプログラミングのブロックにも関与することが示されています。したがって、p53の不活性化により、リプログラミングの効率が向上し、多能性幹細胞が誘導されます。
細胞老化の減速物質製品ラインナップ
幹細胞研究でにおいては、老化の調節因子として以下の2つの化合物が一般に用いられています。
品名/構造式をクリックするとメーカーサイトでの製品の詳細、商品コードをクリックすると各製品の価格表をご覧いただけます。
| 品名/構造式 | 商品コード | CAS番号 | 化学式 | M.W. | 純度 | 標的因子 |
|---|---|---|---|---|---|---|
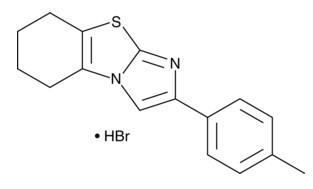
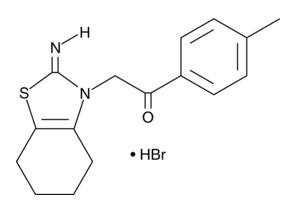
| Cyclic Pifithrin-α (hydrobromide)
|
14748 | 511296-88-1 | C16H16N2S・HBr | 349.3 | ≧95% | p53 |
| Pifithrin-α
|
13326 | 63208-82-2 | C16H18N2OS・HBr | 367.3 | ≧95% |
追加しました。
代謝の減速物質
MEMO 代謝の減速物質とは
オートファジーは、細胞内成分の分解とリサイクルを通じて細胞の恒常性を維持する代謝メカニズムであり、自己複製、分化、老化、アポトーシスを制御します。しかし、これらはすべてタンパク質の代謝回転とリソソームを介するオルガネラ分解の厳密な制御を要します。幹細胞の自己再生および分化中の活性化において、神経幹細胞は、エネルギー消費過程で細胞骨格形状の再構築が必要となります。これは、細胞成分のリサイクル、転写因子、接着分子、または分泌因子一式のリサイクルによるエネルギーを供給するオートファジーの能力に支持されています。オートファジー活性は、多能性幹細胞の発生と維持の間において高レベルで存在します。
多能性幹細胞の生成には、リプログラミング過程において、ミトコンドリアの酸化的リン酸化から解糖へのmTOR調節代謝スイッチが必要となります。mTORは、Sox2によって多能性幹細胞生成の初期段階でリプログラミングが起こるのに必要となる下方制御を受けることが示されています。
SMER28は、オートファゴソームの合成を増加させ、オートファジーの基質の排除を向上させてオートファジーを誘導します。マウス線維芽細胞を神経幹細胞様細胞にリプログラムするために、神経リプログラミング用カクテルのコンポーネントとして用いられます。
代謝の減速物質製品ラインナップ
品名/構造式をクリックするとメーカーサイトでの製品の詳細、商品コードをクリックすると各製品の価格表をご覧いただけます。
| 品名/構造式 | 商品コード | CAS番号 | 化学式 | M.W. | 純度 | 標的因子 |
|---|---|---|---|---|---|---|
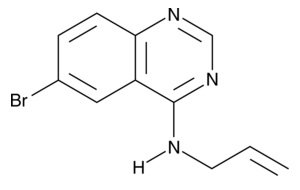
| SMER28
|
17768 | 307538-42-7 | C11H10BrN3 | 264.1 | ≧98% | autophagy |
追加しました。
製品情報は掲載時点のものですが、価格表内の価格については随時最新のものに更新されます。お問い合わせいただくタイミングにより製品情報・価格などは変更されている場合があります。
表示価格に、消費税等は含まれていません。一部価格が予告なく変更される場合がありますので、あらかじめご了承下さい。